
Knoglevæv, ben, benvæv, det specifikke støttevæv, hvoraf hvirveldyrenes knogler er opbygget. Som andre binde- og støttevæv, fx bruskvæv, består knoglevæv af celler lejret i en intercellulær matrix, knoglematrix, der giver vævet dets specifikke mekaniske egenskaber.
knoglevæv

Knoglevæv. Stadier i en bruskpræformeret rørknogles udvikling. 1 Knoglen anlægges i hyalin brusk uden blodkar. Senere udvikler celler i bindevævet omkring knoglens skaft, diafysen, sig til osteoblaster, som danner et tyndt hylster af knoglevæv. Herved forringes den underliggende brusks ernæring, og bruskcellerne dør og efterlader en labyrint af hulrum (2). Disse hulrum invaderes af karførende væv, hvori nogle af cellerne udvikler sig til osteoblaster (3), der begynder at danne knoglevæv. 4 I de tykke partier i knoglens ender, epifyserne, begynder noget senere en tilsvarende proces. Knoglevævsdannelsen i diafysen og epifysen mødes efterhånden omkring en smal bruskplade, epifyseskiven, hvori brusken fortsat vokser, men omdannes til knoglevæv i takt med tykkelsesvæksten. Hos udvoksede fugle og pattedyr omdannes den sidste rest af epifyseskiven til knoglevæv, og længdevæksten kan aldrig genoptages.
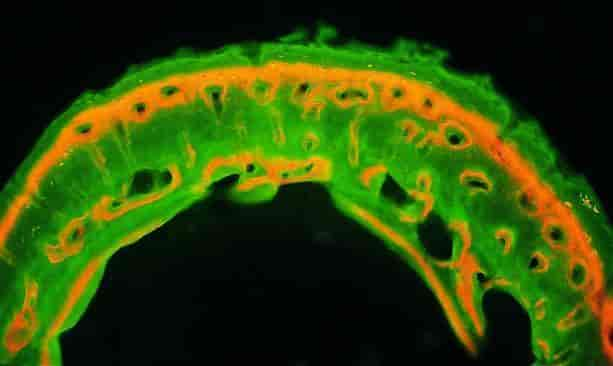
Knoglevæv. Nogle stoffer bindes til knoglevæv under dannelse, bl.a. alizarin (kraprødt). I små doser er stoffet ugiftigt, og indtages det med føden, kan det gennem en periode farve det nydannede knoglevæv. Her ses et tværsnit af skinnebenet fra en rotte. De knoglelameller, der dannedes under alizarinfodringen, fremtræder intenst rødfarvede, mens det øvrige knoglevæv pga. belysningen synes grønligt. Farvefænomenet blev beskrevet i 1736 af en ung læge efter en middag hos en ven, der fremstillede tekstilfarver, herunder kraprødt. Planteaffaldet benyttedes som foder, bl.a. til den gris, der havde udgjort måltidet. Denne opdagelse blev begyndelsen på udviklingen af en metode til studiet af knoglers vækst, som anvendes den dag i dag.
Sammensætning og knogledannelse
Knoglematrix er sammensat af organisk og uorganisk (mineralsk) materiale (primært calcium), der udgør ca. 25 % hhv. 75 % af knoglevævets tørvægt. Langt størstedelen, ca. 90 %, af det organiske materiale udgøres af det trådformede, trækstærke protein kollagen, der dannes (syntetiseres) af de specielle knogledannende celler, osteoblaster (undertiden kaldet bendannere), der også udskiller de enzymer, bl.a. alkalisk fosfatase, som fremkalder udfældning af calciumfosfat i form af hydroxyapatit i den organiske matrix kort efter dens dannelse. Kollagensyntesen lukkes derefter ned, og cellerne overgår i en stofskiftemæssig lavaktiv tilstand som osteocytter, der lever "indstøbt" i lakuner i den mineraliserede matrix, indbyrdes forbundne gennem et net af tynde udløbere. Da den mineraliserede matrix er ueftergivelig, må al vækst af knoglevæv ske ved lagvis tilvækst på overflader, såkaldt appositionel vækst, hvorved knoglevæv får en lamellær indre struktur.
Kompakt knoglevæv
Knoglevæv optræder i to hovedtyper, kompakt og spongiøst knoglevæv. Førstnævnte fremtræder som en kompakt masse, hvori findes mikroskopiske (ca. 50 μm brede) karførende kanaler, Haverske kanaler, samt Volkmannske kanaler, der tværforbinder de Haverske. En Haversk kanal er omgivet af en cylinder af koncentriske knoglelameller, tilsammen betegnet et osteon eller et Haversk lamelsystem; det er knap 0,5 mm i diameter og 15-20 mm langt og er orienteret på langs af de fremherskende retninger for tryk- og trækbelastning af knoglen. Imellem osteonerne findes interstitielle lameller, der repræsenterer rester af nedbrudte osteoner, og som opstår ved knoglevævets konstante omdannelse (remodellering). En knogles overfladiske lag består altid af kompakt knoglevæv, om end dette i visse knogler er ganske tyndt. Yderst er knoglen omgivet af en tæt bindevævshinde, periost, undtagen på flader, hvor knoglen danner ledforbindelser med andre knogler.
Spongiøst knoglevæv
Det spongiøse knoglevæv er opbygget som et tredimensionalt netværk af tynde, indbyrdes sammenhængende bjælker af knoglevæv, trabekler, omgivet af knoglemarv. Trabeklerne indeholder ikke kar, og deres tykkelse er under ca. 250 μm, hvilket er grænsen for ernæring af trabeklernes osteocytter ved diffusion fra den omliggende knoglemarv. Det spongiøse knoglevævs trabekler er, ligesom osteonerne, orienteret svarende til de fremherskende tryk- og trækbelastninger af knoglen.
Tryk- og trækbelastning
Knoglevæv er et kompositmateriale og kan, ligesom jernbeton og glasfiberarmeret plastik, tåle både store trykbelastninger (ca. 20.000 N/cm2) og trækbelastninger (ca. 14.000 N/cm2). Materialet er stift, og stivheden tiltager med alderen. Hos den voksne, hvor spongiosatrabeklerne tåler den største deformering, nås brudgrænsen ved 1-5 % deformation. Visse knogler eller dele heraf er overvejende opbygget af tykt, kompakt knoglevæv, fx de lange knoglers skafter, der pga. rørkonstruktionen opnår stor styrke mod bøjning og torsion med mindst muligt materialeforbrug. Andre knogler, især de, der mest er udsat for trykbelastninger, fx hvirvellegemerne, er overvejende opbygget af spongiøst knoglevæv.
Remodellering og omsætning
Under en organismes vækst forbliver de enkelte knoglers proportioner i hovedtræk konstante, mens den absolutte størrelse tiltager. Derfor må der sideløbende med nydannelse af knoglevæv finde nedbrydning sted; fx må der under hjernens vækst fjernes knoglevæv på hjernekassens inderside, mens nyt knoglevæv dannes på dens yderside. Denne nedbrydning varetages af en særlig celletype, osteoclaster (undertiden kaldet benædere), der fæstner sig til knoglevævet og udskiller syre og hydrolytiske (lysosomale) enzymer, hvorved såvel den uorganiske som den organiske del af knoglematrix nedbrydes.
Nedbrydning og opbygning af knoglevæv finder sted gennem hele livet. Omsætningen er i barnealderen i størrelsesordenen 200 % årlig, hos den voksne blot nogle få procent. Ved nedbrydning (resorption) af et osteon danner en gruppe osteoclaster et "borehoved", der udborer osteonet på langs. Lige efter de avancerende osteoclaster følger osteoblaster, der opbygger nyt knoglevæv på resorptionskanalens vægge. Tilsvarende vil der på spongiosatrabeklerne på samme tid findes osteoclaster og osteoblaster, der udøver deres modsatrettede virksomhed. Ved ændrede belastningsforhold af en knogle vil spongiosastrukturen på kort tid (i løbet af måneder) ombygges svarende til de nye forhold. Balancen mellem opbygning og nedbrydning samt knoglevævets mineraliseringsgrad påvirkes af en lang række faktorer i et kompliceret samspil. Navnlig hormonelle og mekaniske, men også ernæringsforhold spiller ind, især kostens indhold af calcium og D-vitamin (se calcium-stofskiftet, engelsk syge og knogleskørhed).
Osteoblaster stimuleres til at danne knoglevæv af hormonerne calcitonin, somatotropin (væksthormon) samt kønshormon, mens osteoclasternes knoglenedbrydning stimuleres af parathyroideahormon. Af væsentlig betydning for denne balance er den mekaniske belastning af knoglevævet. Ved sengeleje eller i vægtløs tilstand (under astronauters længerevarende ophold i rummet) kan knoglerne tabe op mod 1 % af deres mineralindhold pr. uge. Omvendt vil øget belastning som fx sportstræning øge både mængden af knoglevæv og dets mineraliseringsgrad.
Med alderen sker der naturligt en reduktion af mængden af knoglevæv i knoglerne, idet hulrummene bliver større (såkaldt osteoporose), ligesom mineraliseringsgraden kan aftage (halisterese). Hos kvinder medfører den nedsatte produktion af østrogen efter klimakteriet, at balancen hurtigere end hos mænd forskydes mod nedbrydning.
Knoglevævets udviklingshistorie
Knoglevæv findes kun hos hvirveldyr, hvor det optræder i to forskellige udgaver og med to forskellige dannelsesmekanismer. Den tidligste udvikledes for 500 mio. år siden som knogleplader dannet i bindevævet (dermis) under overfladeepithelet hos visse fiskeformer; ekstremt udviklet hos Devonhavets panserhajer, se placodermer. Denne knogledannelse er kendetegnet ved, at celler i hudens bindevæv direkte differentierer til osteoblaster, således at knoglevævet opstår uden et forudgående bruskmellemstadium, og knoglerne betegnes bindevævspræformerede. Denne forbeningsmåde er evolutionært videreført og optræder i forskellige varianter hos alle senere hvirveldyrklasser; hos pattedyrene bl.a. ved dannelsen af de flade knogler i hjernekassen samt underkæbeknoglen.
Senere, tidligst i Devon, opstod en anden type knogler hos de tidlige hvirveldyr, hvis indre bruskskelet blev "byttet ud" med et knogleskelet ved en proces, hvor knoglen først anlægges som en bruskknogle, der derefter nedbrydes indefra under samtidig opbygning af knoglevæv, der efterhånden erstatter bruskvævet under bevarelse af knoglens ydre form. Sådanne knogler betegnes erstatningsknogler eller bruskpræformerede knogler, og de udgør størstedelen af de højerestående hvirveldyrs skelet. Et rent bruskskelet findes bevaret hos bruskfiskene, bl.a. hajer og rokker, men hos de evolutionært senere hvirveldyr, inkl. mennesket, anlægges skelettet i fosterlivet stadig som et bruskskelet, og først ved vækstens afslutning bliver de sidste rester af bruskskelettet erstattet af knoglevæv, se også skelet.
Knoglevæv dannes undertiden som afstivende element andre steder i legemet. I penis dannes hos mange pattedyr, bl.a. ulve, hunde, bjørne og sæler, en knogle, baculum, i skillevæggen mellem svulmelegemerne. Nært beslægtet med knoglevæv er det bindevævspræformerede tandben, dentin, se tænder.
Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.