De enkelte muskler er sammensat af type I- og II-fibre i et blandingsforhold, der afspejler de normale funktionskrav til musklen, men forholdet er påvirket af individuelle, genetisk bestemte variationer, som gør nogle mennesker egnede til udholdenhedskrævende muskelarbejde (fx maratonløb), andre til kortvarigt, hurtigt og kraftfuldt muskelarbejde (fx spring, sprint og kampsport).
Effekten af udholdenhedstræning beror især på øgning af antallet af mitokondrier i musklernes type I-fibre og af tætheden af kapillærer mellem fibrene. Ved styrketræning, dvs. træning med høj belastning, øges også antallet af myofibriller, især i type II-fibrene, navnlig hvis tempoet er højt.
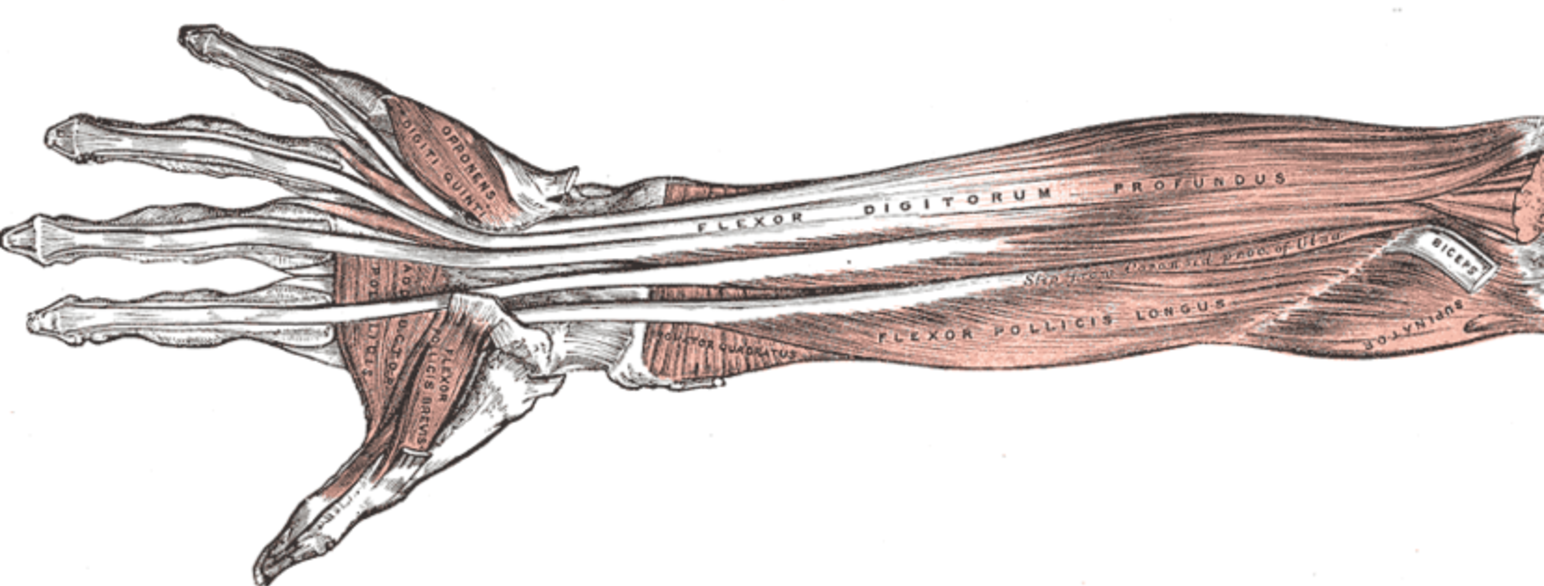
En enkelt nervetråd danner vha. forgreninger motoriske endeplader på flere muskelfibre, der alle er af den samme type, og som kontraherer sig samtidig; tilsammen en motor unit ('motorisk enhed'). Antallet af muskelfibre i en motor unit varierer meget. I præcist styrede muskler, fx øjenmusklerne, er antallet knap ti; i fingrenes små muskler, der bruges til præcisionsbevægelser, er antallet 100-300; i de kraftfulde, men grove hofte- og benmuskler op til 2000.
Inden for den enkelte muskel danner type I-fibrene mindre motor units end type II-fibrene, der først aktiveres, når behovet for muskelkraft overstiger en vis størrelse. I en muskel, der arbejder konstant gennem længere tid, vil de enkelte motor units skifte mellem arbejds- og hvileperioder.
En væsentlig effekt af idrætstræning består i optimering af antal og type af motor units, der aktiveres af centralnervesystemet under udførelsen af de med arbejdet forbundne bevægelser, således at en bevægelse kan forbedres og udføres næsten refleksmæssigt.
Et afgørende element i denne tilpasning er de oplysninger, der tilgår centralnervesystemet fra musklernes specielle sanseorganer, muskel- og senetene. Muskeltenene er indlejret i selve musklen og registrerer dels musklens længde, dels den hastighed, hvormed længden ændres under bevægelsen. Senetenene er indlejret i de sener, der overfører muskeltrækket, og registrerer musklens spænding, tonus. Disse sanseoplysninger sammenholdes i centralnervesystemet med oplysninger fra sanseorganer i ledkapsler og ledbånd og kommer til bevidsthed som stillingssansen, der (selv når man har lukkede øjne) giver præcis viden om legemets stilling eller rumlige bevægelse.
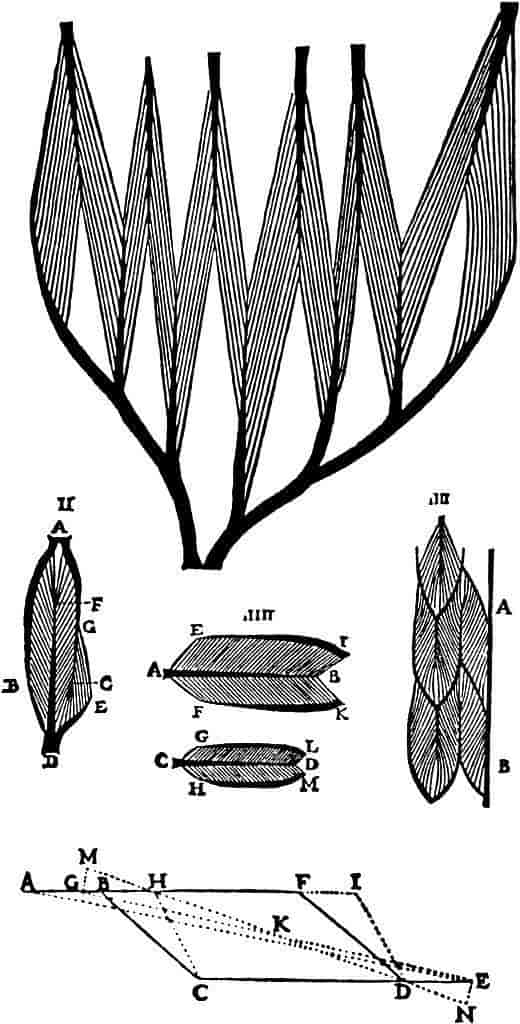
En skeletmuskels kraftudvikling og dens arbejdsydelse, effekt, afhænger af flere faktorer, især af muskelcellernes arrangement i musklen. En muskelcelle kan udvikle en kraft, der er ligefrem proportional med tværsnitsarealet af dens myofibriller. Er fibren lang, kan den forkorte sig mere, men kraften er den samme uanset længden. Mange store muskler er opbygget af et stort antal korte fibre, der er tilhæftet seneblade beliggende inde i musklen, de såkaldte pennate muskler. I disse er det samlede tværsnitsareal af muskelfibre større, end hvis musklen var opbygget af lange, parallelle fibre. Musklen får på denne måde et stort såkaldt fysiologisk tværsnit og kan udvikle en tilsvarende større kraft.
Kraftudviklingen (spændingen) er alt andet lige størst, når musklen kontraherer sig nær dens normale hvilelængde uden at forkortes, dvs. ved såkaldt isometrisk arbejde, modsat isotonisk arbejde, hvor musklen kontraheres under fast kraftudvikling. Er længden større, aftager spændingen, da aktin- og myosinfilamenterne overlapper mindre; ved strækning til omkring 160% af hvilelængden er kraften lig nul. Ved forkortning til under 75% af hvilelængden aftager kraften hurtigt og falder til nul ved 40% forkortning, da aktinfilamenterne er trukket helt ind mellem myosinfilamenterne og støder ende mod ende.
Når musklen arbejder under samtidig forkortning, såkaldt koncentrisk arbejde, aftager kraften, hvis kontraktionshastigheden øges. Effektydelsen, udtrykt ved produktet af kraft og hastighed, er maksimal, når musklen arbejder omkring sin hvilelængde og kontraherer sig med ca. 1/3 af den maksimale hastighed; det er vigtigt at tage dette forhold i betragtning ved fx maksimering af arbejdsydelsen i cykelløb.
Muskler arbejder ofte under samtidig forlængelse, såkaldt excentrisk arbejde, som det fx finder sted ved opbremsning under almindelig gang, især ned ad bakke. Ved store opbremsninger, fx et nedspring, hvor der skal absorberes en stor energimængde af de excentrisk arbejdende muskler, er muskelskader hyppige, især fordi dette arbejde stiller store krav til nervesystemets præcise koordinering af aktiviteten af motor units.
Nytteværdien ved muskelarbejde, dvs. den del af den tilførte kemiske energimængde, der kan genvindes som mekanisk arbejde, varierer stærkt og afhænger af arbejdets art og træningsgraden, men ligger for mennesket i de fleste situationer fra 15% til højst 25%. De ikke udnyttede 75% til 85% af den tilførte energi forlader musklen som varme.



Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.