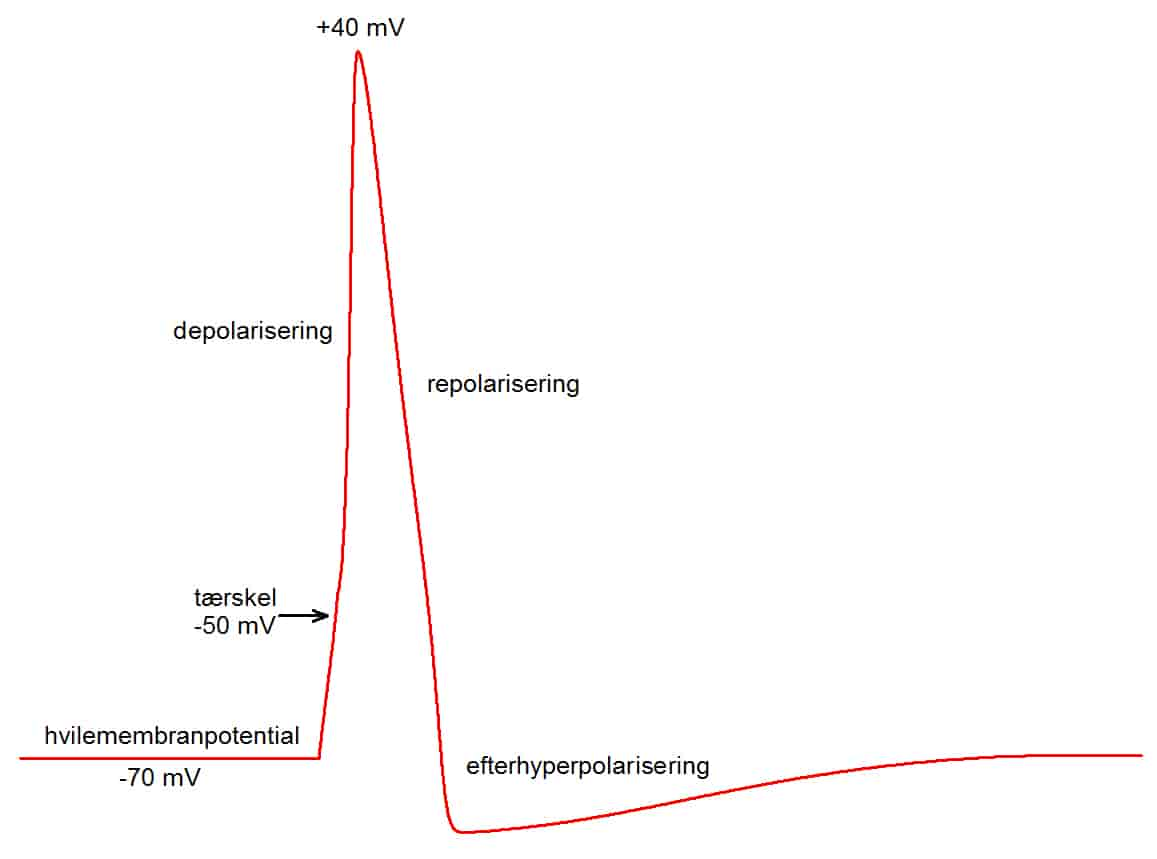
Et eksempel på kemisk stimulation er synaptisk transmission, som er den generelle måde, hvorpå nerveceller overfører signaler til hinanden. Nerveceller danner små kontaktpunkter kaldet synapser. I synapser overføres signaler sædvanligvis kun en vej. Hvis afsendercellen affyrer et aktionspotential, frigives signalstof kaldet en transmitter eller neurotransmitter til det lille rum mellem de to nerveceller i synapsen. Når neurotransmitter-molekylerne er frigivet, sætter de sig på specialiserede molekyler (receptorer) på modtagercellens membran. Receptorerne fungerer, samtidig med at de binder neurotransmitteren, som ionkanaler, der åbnes af bindingen af transmitteren til receptoren. Hvis receptor/ionkanalen tillader strøm af f.eks. Na+ ind i modtagercellen, så depolariseres modtagercellen, og hvis det er en receptor/ionkanal, der tillader f.eks. K+ at strømme ud af cellen, så hyperpolariseres modtagercellen. Neurotransmitteren fjernes hurtigt ved nedbrydning eller ved at blive optaget af de omgivende celler, så den synaptiske transmission varer sædvanligvis kun nogle få millisekunder
Denne form for kemisk signaloverførsel kaldes synaptisk transmission. Depolariseringen kaldes et excitatiorisk synaptisk potential (EPSP) og hyperpolariseringen kaldes et inhibitorisk synaptisk potential (IPSP). Synaptiske potentialer er i reglen små (1 – 10 mV), men hvis flere EPSP'er forekommer samtidig, så summeres de, og derved skabes en depolarisering, der er stor nok til at tærsklen for et aktionspotential bliver nået. Det betyder, at et aktionspotential i en nervecelle ikke nødvendigvis udløser et aktionspotential i en anden nervecelle, som den er i kontakt med, men aktionspotentialet kan øge sandsynligheden for, at modtagercellen affyrer et aktionspotential. IPSP'er fjerner membranpotentialet fra tærskelværdien, og derfor fungerer disse synaptiske potentialer som hæmmere. De mindsker sandsynligheden for, at der opstår et aktionspotential.
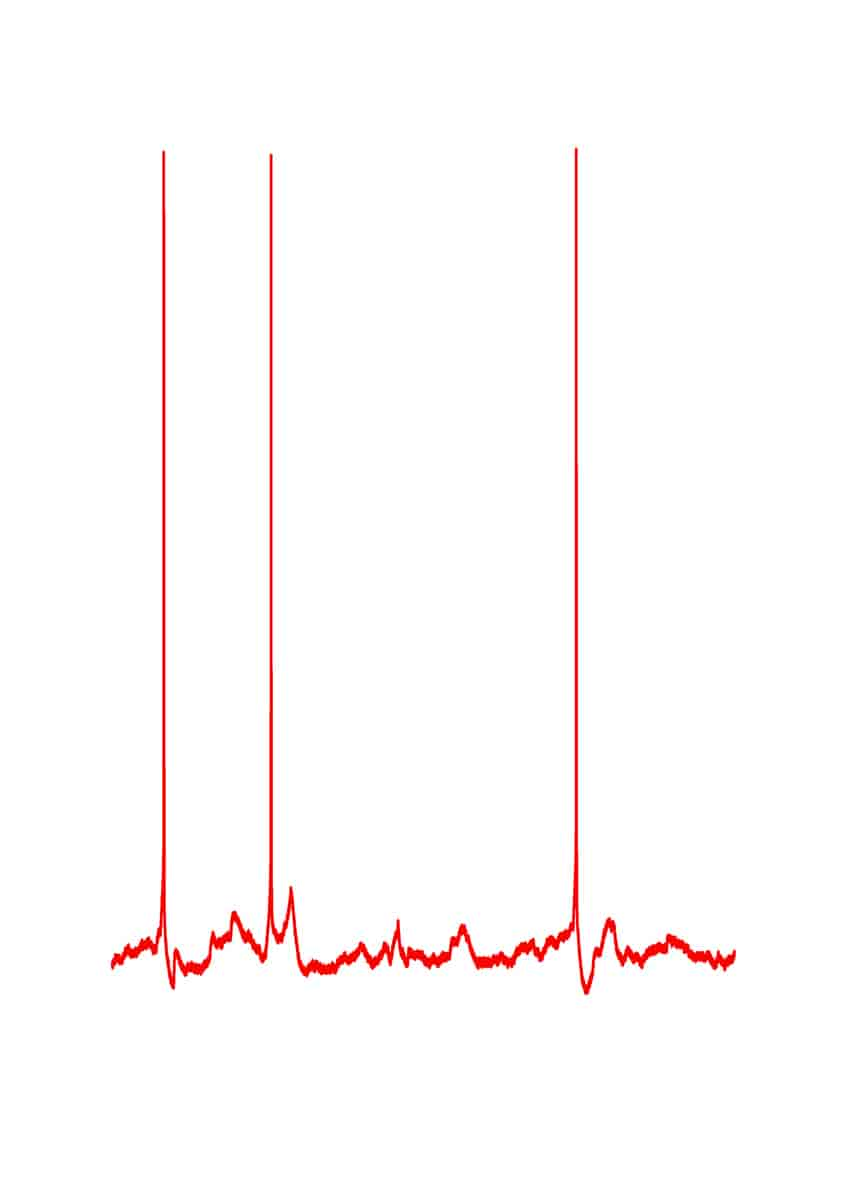
Eksperimentelt kan nerveceller isoleres, og så har de et stabilt hvilemembranpotential, men når cellerne sidder i netværk af nerveceller, så bombarderes de konstant af synaptisk aktivitet, og så fluktuerer membranpotentialet. Et eksempel herpå ses i figuren.


Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.